

氮肥在水草的栽培上,
向來是個很熱門的話題,
最熱門的要算是透過缺氮的方式來促使水草變紅,
以及氨氮(NH4)或硝酸氮(NO3)哪一個比較會導致爆藻。
水草所需的氮肥來源也是個很被人提及的議題,
例如許多研究發現水草以氨氮當作氮肥優先吸收型態,
可是水草對於氨氮的耐受濃度卻又比硝酸氮來得低。
以金魚藻(Ceratophyllum demersum)為例,
我們在水草偏好哪一種氮肥?一文中提到了,
金魚藻的成長在水體中氨氮達 5 ppm 時受到了抑制,
但是在硝酸氮 350 ppm 的溶液中還能吸收成長。
這是個非常有趣的現象,
許多水草既然偏愛氨氮,
那為何氨氮對水草的毒害或抑制濃度也不高?
這似乎是很不符合邏輯的現象,
況且水草對其他營養素的忍受濃度範圍,
也似乎都比氨氮來得高很多。
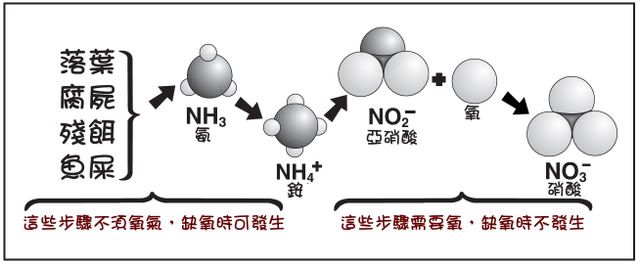
我們現在已經知道了,
水草對於氨氮可以直接的吸收利用,
而對於硝酸氮則必須先透過硝酸還原酶(Nitrate reductase)的作用,
還原成亞硝酸(NO2)並進一步還原成氨氮(NH4)以後,
才能進一步合成氨基酸和蛋白質,
而將硝酸還原成氨氮的過程是需要消耗能量的,
根據 Diana Walstad 女士的推測,
在理論上這個過程需要消耗 83 Kcal/mol 的能量。
水草從頂芽至根部的外均有氧氣,不但防止了許多物質的毒害,也都有硝化細菌附著,將氨氮變成了硝酸。
換個角度來看,
水草如果不想浪費能量的話,
那麼直接吸收氨氮就好了,
可是如果水草需要運用硝酸氮做為營養的來源時,
硝酸還原酶的活性就會比較高,
也就是會消耗較多的能量來獲取氮元素。
依照一般人的邏輯來看,
水草會對於氮肥的吸收會以氨氮為主,
必要的時候才會消耗能量來吸收硝酸。
其實根據陸生植物的研究發現,
成長緩慢的植物如樹木和矮灌木,
硝酸還原酶的作用或活性主要集中在根部,
也就是說先在根部將硝酸還原成氨氮,
再運送至植物的各個部位去使用。
而快速生長的作物,
則傾向先將大部分的硝酸運送至頂芽,
然後才進行還原的作用,
也就是說頂芽的硝酸還原酶活性是高於根部的。
此外當外在的硝酸濃度增高時,
頂芽的硝酸還原酶活力增加也大於根部,
所以一般認為植物頂芽的硝酸還原酶活性,
和抵達頂芽的硝酸濃度有關。
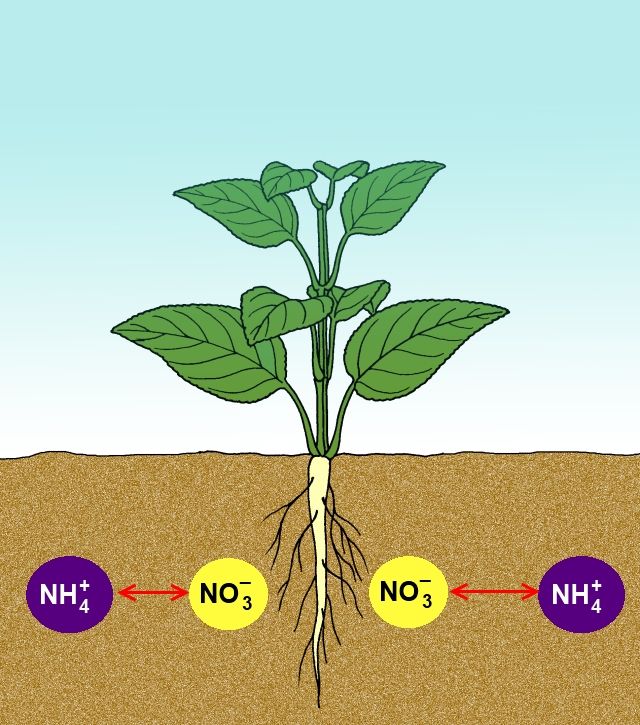
那麼水草呢?
水草和陸生植物不一樣,
在水下的生活環境中,
不論是在水體或底床,
周遭都有相當豐富的氮元素,
例如含氧量較高水體中以硝酸氮為主,
在嚴重缺氧的底床環境則以氨氮為主,
除非是在溶氧量較高的粗粒底床中。
所以按照常理來判斷,
著根於底泥和水流緩慢溪流的水草,
只會從頂芽吸收明顯份量的硝酸;
至於漂浮性水草和著根於高溶氧量底床的水草,
不論根部或頂芽都很可能會吸收硝酸。
水草根系會向周遭釋出氧氣,有助於硝化作用的進行,也就是說將氨氮轉變成了硝酸。
此外我們也能夠預期,
水草體內硝酸還原酶的活性,
會因外在環境硝酸濃度增加而增高;
而當外在環境存在著氨氮時,
水草的硝酸環原酶也會受到干擾。
若從環境的角度來看,
著根生長於缺氧底床的,
頂芽的硝酸還原酶活性應該會高於根部的活性,
因為位於缺氧底床的根部可以直接吸收氨氮,
理論上不太需像頂芽一樣消耗能量吸收硝酸。
不過我們常說,
不可以將陸生植物的研究直接套用至水草,
理論上的推想也不必然等於事實,
凡事都要實驗求證才能知道。
丹麥的學者 Cedergreen 等人在 2003 年發表了一篇研究,
針對水草頂芽和根部硝酸還原酶進行了實驗,
讓我們有機會更深入了解水草對氮肥的吸收。
研究人員針對十二種水草進行了頂芽和根部硝酸還原酶進行了活性測量,
這十二種水草包括了漂浮性的水草如浮萍(Lemna gibba)、浮葉性的水草如黃花萍蓬草(Nuphar lutea)和沒有根的金魚藻等等。
丹麥的研究人員發現,
剛從野外採集回來的水草中,
只有大苦水芹(Cardamine amara)頂芽的硝酸還原酶活性還算高,
其他的十一種水草不論是根部或頂芽之硝酸還原酶活性都很低。
浮萍在添加硝酸以後,根部的硝酸還原酶活性增加遠大於頂芽。

不過丹麥的學者發現,
水草的硝酸還原酶活性是可以被誘發的,
他們加入了 500 μM(31 ppm)的硝酸以後,
受測的五種水草之硝酸還原酶大幅的提高 2 至 17 倍。
另外在十七株測量硝酸還原酶的水草當中,
有十一株的根部活性是高於頂芽的。
這也就是說,
在水體內添加了硝酸以後,
根部和頂芽的硝酸還原酶或性都會增加,
而且根系甚至超過了頂芽。
例外的是依樂藻(Elodea canadensis),
依樂藻在加入硝酸以後,
雖然頂芽的硝酸還原酶或性增加了,
根部的活性反而是降低的。
這個結果雖然說明了水草根系在硝酸還原所扮演的重要角色,
也讓丹麥的研究人員很出乎意料。
我們也不禁感到納悶,
這個實驗結果所意涵的,
到底是水草會將硝酸運送至根部再進行還原作用呢?
或者水草進行硝酸還原作用的部位主要是位於根部?
而我們如果想要讓水草更積極的吸收硝酸,
是否要如研究一樣加重濃度至 31 ppm 才更能促進硝酸還原酶的活性?
影響到硝酸還原酶活性的因素很多,
例如氨氮的存在、溫度、光照和碳元素,
都會影響到植物硝酸還原酶的活性。
以溫度對硝酸還原酶的影響來說,
當北極植物的根區溫度由 10 度上升至 20 度時,
硝酸還原酶的活性最大可增加一倍。
氨氮的存在可說是很令人感到興趣的。
的確有不少研究發現氨氮的存在會影響到水草對硝酸的吸收。
聚藻(Myriophyllum spicatum)在加入了 2 μM(0.036 ppm)的氨氮濃度後,
對於 24 μM(1.49 ppm)硝酸的吸收就會受到阻礙;
浮萍要在氨氮濃度小於 10 μM(0.18 ppm)才會開始吸收硝酸。
水草實驗中的氨氮(NH4)降低,到底表面硝化細菌的作用,或者是真的被水草吸收,是個耐人尋味的問題。
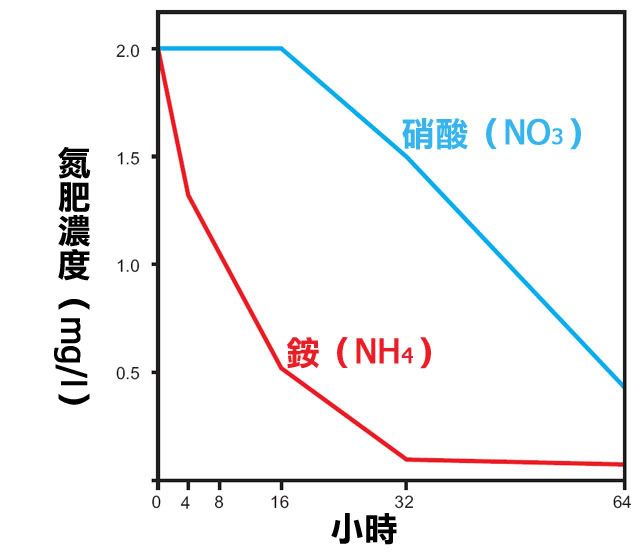
這又不禁令我們想起了 Ozimek 在 1990 年針對美洲小水蘊草(Elodea nuttallii)的研究,
在同樣是 2 ppm 氨氮和硝酸濃度的條件下,
氨氮的濃度下降比較快,
對此 Diana Walstad 女士認為水草偏好吸收氨氮。
可是 Tom Barr 卻以另一種角度來解讀,
他認為水草在氨氮小於 0.5 ppm 時,
水草對於氨氮的吸收將減緩,
對於硝酸的吸收將增快;
而當氨氮濃度低於 0.1 ppm 時,
水草將不再吸收氨氮了。
所以 Tom Barr 認為在一般的水草缸中,
硝酸才是水草氮肥的主要來源,
因為一般水草缸的氨氮濃度絕少超過 0.1 ppm 。
換句話說,
一般研究所使用的氨氮濃度,
很少出現在我們的水族缸,
因為那些濃度對我們的水族動物大都已經是毒害的濃度了。
所以造成抑制硝酸吸收的氨氮濃度在我們的水族缸是很難發生的。
另一個議題是水草根部的硝化做用。
水草莖葉表面的硝化作用非常的活躍,
水草能吸收得到氨氮的機會本來就不高。
我們在水草對脫氮作用的影響(上):根系篇提到過,
水草的根部不分晝夜都會向周遭釋出氧氣,
提供硝化細菌生長所需。
丹麥的學者 Richard-Petersen 在 1997 年的研究發現,
以 24 小時光照和 24 小時黑暗的實驗方式,
測得水草根部在經過 24 小時的黑暗期後,
周圍仍舊有氧氣存在,
也就是說足以維持硝化作用繼續進行。
使用泥土的天然水草缸,水草根部吸收到氨氮的機會,比使用粗粒底砂的水草缸來得高。

丹麥的研究發現在水體加入硝酸後,
大部分的水草根部硝酸還原酶活性高於頂芽。
從另一個角度來看,
硝酸能夠滲透至底床內由根部吸收,
可是氨氮則大都停留在底床內,
因此根系除了要處理來自週遭硝化作用所產生的硝酸以外,
也要應付人為的硝酸添加。
這似乎也可以用來說明為何根系的硝酸還原酶活性較高的原因。
當然了,
我們也可以從質量增加的角度來解讀,
也就是說根系的成長比頂芽還要快,
所以需要更多的氮元素,
並且反映在硝酸還原酶的活性上。
無論如何,
大部分水草缸之水體氨氮濃度並不高,
底床除非是天然水草缸使用底泥,
再說底材也是以粗粒的底砂(相較於泥土)為主,
底床的溶氧量並不會過低,
也就是說氨氮的濃度不致累積過高。
這也不禁令人懷疑,
既然水草的根系周圍並非是缺氧狀態,
況且還是硝化作用很活躍的區域,
那麼水草還有多少機會可以吸收得到氨氮?
向來是個很熱門的話題,
最熱門的要算是透過缺氮的方式來促使水草變紅,
以及氨氮(NH4)或硝酸氮(NO3)哪一個比較會導致爆藻。
水草所需的氮肥來源也是個很被人提及的議題,
例如許多研究發現水草以氨氮當作氮肥優先吸收型態,
可是水草對於氨氮的耐受濃度卻又比硝酸氮來得低。
以金魚藻(Ceratophyllum demersum)為例,
我們在水草偏好哪一種氮肥?一文中提到了,
金魚藻的成長在水體中氨氮達 5 ppm 時受到了抑制,
但是在硝酸氮 350 ppm 的溶液中還能吸收成長。
這是個非常有趣的現象,
許多水草既然偏愛氨氮,
那為何氨氮對水草的毒害或抑制濃度也不高?
這似乎是很不符合邏輯的現象,
況且水草對其他營養素的忍受濃度範圍,
也似乎都比氨氮來得高很多。
我們現在已經知道了,
水草對於氨氮可以直接的吸收利用,
而對於硝酸氮則必須先透過硝酸還原酶(Nitrate reductase)的作用,
還原成亞硝酸(NO2)並進一步還原成氨氮(NH4)以後,
才能進一步合成氨基酸和蛋白質,
而將硝酸還原成氨氮的過程是需要消耗能量的,
根據 Diana Walstad 女士的推測,
在理論上這個過程需要消耗 83 Kcal/mol 的能量。
水草從頂芽至根部的外均有氧氣,不但防止了許多物質的毒害,也都有硝化細菌附著,將氨氮變成了硝酸。
換個角度來看,
水草如果不想浪費能量的話,
那麼直接吸收氨氮就好了,
可是如果水草需要運用硝酸氮做為營養的來源時,
硝酸還原酶的活性就會比較高,
也就是會消耗較多的能量來獲取氮元素。
依照一般人的邏輯來看,
水草會對於氮肥的吸收會以氨氮為主,
必要的時候才會消耗能量來吸收硝酸。
其實根據陸生植物的研究發現,
成長緩慢的植物如樹木和矮灌木,
硝酸還原酶的作用或活性主要集中在根部,
也就是說先在根部將硝酸還原成氨氮,
再運送至植物的各個部位去使用。
而快速生長的作物,
則傾向先將大部分的硝酸運送至頂芽,
然後才進行還原的作用,
也就是說頂芽的硝酸還原酶活性是高於根部的。
此外當外在的硝酸濃度增高時,
頂芽的硝酸還原酶活力增加也大於根部,
所以一般認為植物頂芽的硝酸還原酶活性,
和抵達頂芽的硝酸濃度有關。
那麼水草呢?
水草和陸生植物不一樣,
在水下的生活環境中,
不論是在水體或底床,
周遭都有相當豐富的氮元素,
例如含氧量較高水體中以硝酸氮為主,
在嚴重缺氧的底床環境則以氨氮為主,
除非是在溶氧量較高的粗粒底床中。
所以按照常理來判斷,
著根於底泥和水流緩慢溪流的水草,
只會從頂芽吸收明顯份量的硝酸;
至於漂浮性水草和著根於高溶氧量底床的水草,
不論根部或頂芽都很可能會吸收硝酸。
水草根系會向周遭釋出氧氣,有助於硝化作用的進行,也就是說將氨氮轉變成了硝酸。
此外我們也能夠預期,
水草體內硝酸還原酶的活性,
會因外在環境硝酸濃度增加而增高;
而當外在環境存在著氨氮時,
水草的硝酸環原酶也會受到干擾。
若從環境的角度來看,
著根生長於缺氧底床的,
頂芽的硝酸還原酶活性應該會高於根部的活性,
因為位於缺氧底床的根部可以直接吸收氨氮,
理論上不太需像頂芽一樣消耗能量吸收硝酸。
不過我們常說,
不可以將陸生植物的研究直接套用至水草,
理論上的推想也不必然等於事實,
凡事都要實驗求證才能知道。
丹麥的學者 Cedergreen 等人在 2003 年發表了一篇研究,
針對水草頂芽和根部硝酸還原酶進行了實驗,
讓我們有機會更深入了解水草對氮肥的吸收。
研究人員針對十二種水草進行了頂芽和根部硝酸還原酶進行了活性測量,
這十二種水草包括了漂浮性的水草如浮萍(Lemna gibba)、浮葉性的水草如黃花萍蓬草(Nuphar lutea)和沒有根的金魚藻等等。
丹麥的研究人員發現,
剛從野外採集回來的水草中,
只有大苦水芹(Cardamine amara)頂芽的硝酸還原酶活性還算高,
其他的十一種水草不論是根部或頂芽之硝酸還原酶活性都很低。
浮萍在添加硝酸以後,根部的硝酸還原酶活性增加遠大於頂芽。
不過丹麥的學者發現,
水草的硝酸還原酶活性是可以被誘發的,
他們加入了 500 μM(31 ppm)的硝酸以後,
受測的五種水草之硝酸還原酶大幅的提高 2 至 17 倍。
另外在十七株測量硝酸還原酶的水草當中,
有十一株的根部活性是高於頂芽的。
這也就是說,
在水體內添加了硝酸以後,
根部和頂芽的硝酸還原酶或性都會增加,
而且根系甚至超過了頂芽。
例外的是依樂藻(Elodea canadensis),
依樂藻在加入硝酸以後,
雖然頂芽的硝酸還原酶或性增加了,
根部的活性反而是降低的。
這個結果雖然說明了水草根系在硝酸還原所扮演的重要角色,
也讓丹麥的研究人員很出乎意料。
我們也不禁感到納悶,
這個實驗結果所意涵的,
到底是水草會將硝酸運送至根部再進行還原作用呢?
或者水草進行硝酸還原作用的部位主要是位於根部?
而我們如果想要讓水草更積極的吸收硝酸,
是否要如研究一樣加重濃度至 31 ppm 才更能促進硝酸還原酶的活性?
影響到硝酸還原酶活性的因素很多,
例如氨氮的存在、溫度、光照和碳元素,
都會影響到植物硝酸還原酶的活性。
以溫度對硝酸還原酶的影響來說,
當北極植物的根區溫度由 10 度上升至 20 度時,
硝酸還原酶的活性最大可增加一倍。
氨氮的存在可說是很令人感到興趣的。
的確有不少研究發現氨氮的存在會影響到水草對硝酸的吸收。
聚藻(Myriophyllum spicatum)在加入了 2 μM(0.036 ppm)的氨氮濃度後,
對於 24 μM(1.49 ppm)硝酸的吸收就會受到阻礙;
浮萍要在氨氮濃度小於 10 μM(0.18 ppm)才會開始吸收硝酸。
水草實驗中的氨氮(NH4)降低,到底表面硝化細菌的作用,或者是真的被水草吸收,是個耐人尋味的問題。
這又不禁令我們想起了 Ozimek 在 1990 年針對美洲小水蘊草(Elodea nuttallii)的研究,
在同樣是 2 ppm 氨氮和硝酸濃度的條件下,
氨氮的濃度下降比較快,
對此 Diana Walstad 女士認為水草偏好吸收氨氮。
可是 Tom Barr 卻以另一種角度來解讀,
他認為水草在氨氮小於 0.5 ppm 時,
水草對於氨氮的吸收將減緩,
對於硝酸的吸收將增快;
而當氨氮濃度低於 0.1 ppm 時,
水草將不再吸收氨氮了。
所以 Tom Barr 認為在一般的水草缸中,
硝酸才是水草氮肥的主要來源,
因為一般水草缸的氨氮濃度絕少超過 0.1 ppm 。
換句話說,
一般研究所使用的氨氮濃度,
很少出現在我們的水族缸,
因為那些濃度對我們的水族動物大都已經是毒害的濃度了。
所以造成抑制硝酸吸收的氨氮濃度在我們的水族缸是很難發生的。
另一個議題是水草根部的硝化做用。
水草莖葉表面的硝化作用非常的活躍,
水草能吸收得到氨氮的機會本來就不高。
我們在水草對脫氮作用的影響(上):根系篇提到過,
水草的根部不分晝夜都會向周遭釋出氧氣,
提供硝化細菌生長所需。
丹麥的學者 Richard-Petersen 在 1997 年的研究發現,
以 24 小時光照和 24 小時黑暗的實驗方式,
測得水草根部在經過 24 小時的黑暗期後,
周圍仍舊有氧氣存在,
也就是說足以維持硝化作用繼續進行。
使用泥土的天然水草缸,水草根部吸收到氨氮的機會,比使用粗粒底砂的水草缸來得高。
丹麥的研究發現在水體加入硝酸後,
大部分的水草根部硝酸還原酶活性高於頂芽。
從另一個角度來看,
硝酸能夠滲透至底床內由根部吸收,
可是氨氮則大都停留在底床內,
因此根系除了要處理來自週遭硝化作用所產生的硝酸以外,
也要應付人為的硝酸添加。
這似乎也可以用來說明為何根系的硝酸還原酶活性較高的原因。
當然了,
我們也可以從質量增加的角度來解讀,
也就是說根系的成長比頂芽還要快,
所以需要更多的氮元素,
並且反映在硝酸還原酶的活性上。
無論如何,
大部分水草缸之水體氨氮濃度並不高,
底床除非是天然水草缸使用底泥,
再說底材也是以粗粒的底砂(相較於泥土)為主,
底床的溶氧量並不會過低,
也就是說氨氮的濃度不致累積過高。
這也不禁令人懷疑,
既然水草的根系周圍並非是缺氧狀態,
況且還是硝化作用很活躍的區域,
那麼水草還有多少機會可以吸收得到氨氮?

 RSS 訂閱
RSS 訂閱